Transkriptsioonifaktor Gal4
Transkriptsioonifaktor Gal4 (881 aminohapet) koosneb mitmest erinevast domeenist. Põhilised domeenid on:
- aktivatsioonidomeen (aminohapped 750-868) - vajalik transkriptsiooni aktiveerimiseks
- DNA-ga seostumise domeen (aminohapped 14-57) - sisaldab Zn2/Cys6 motiivi, seostub UASG järjestusele promootoralas
- dimerisatsioonidomeen (aminohapped 50-108) - vajalik dimeeri moodustamisel
- Gal80 seostumise domeen (aminohapped 856-869)
Lisaks paikneb valgu N-terminaalses osas tuumalokalisatsiooni signaal ning keskmises regioonis glükoosi inhibitsiooni domeen.
Gal4 aktivatsiooni - ja DNA-ga seostumise domeeni olemasolu tõestati lihtsate deletsioonanalüüsi katsete abil (Joonis 4). Katsete eesmärgiks oli analüüsida erinevate mutantide võimet aktiveerida reportergeeni ekspressiooni. GAL geenide promootorala paigutati lacZ reportergeeni ette ning mõõdeti reportergeeni ekspressiooni.
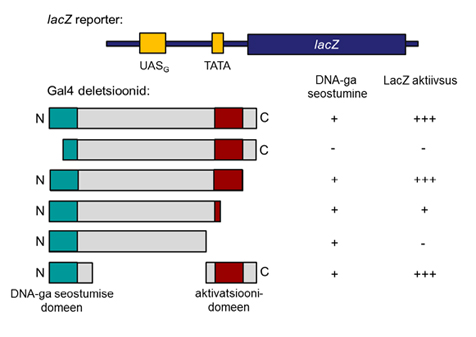
Joonis 4. Gal4 deletsioonanalüüs. Näidatud on kasutatud reportergeeni struktuur, erinevad deletsioonimutandid ning ende võime seostuda DNA-ga ja aktiveerida reportergeeni ekspressiooni.
Gal4 deletsioonanalüüs näitas, et kui eemaldada valgu N-terminaalne domeen, siis selline mutant ei ole enam võimeline seostuma UASG järjestusele. DNA-ga seostumine on eelduseks transkriptsiooni aktivatsioonile. Seega ei aktiveeri selline mutant ka reportergeeni transkriptsiooni. Eemaldades Gal4 C-terminaalse domeeni, kadus mutandil võime aktiveerida reportergeeni transkriptsiooni. Samas säilis DNA-ga seostumine. Valgu keskmise regiooni eemaldamisel säilis mutandil võime seostuda DNA-ga ning aktiveerida transkriptsioon.
Gal4 domeenide paiknemist on uuritud ka domeeni vahetust (domain swapping) kasutades (Joonis 5). Konstrueeriti kolm liitvalku: Gal4 aktivatsioonidomeen (Gal4-AD) liideti E. coli valgu LexA DNA-ga seostumise domeeniga (LexA-BD). Bakteriaalne LexA seostub DNA järjestusele, mida nimetatakse LexA operaator (LexA OP). Teine liitvalk koosnes Gal4 DNA-ga seostumise domeenist (Gal4-BD) ja viirusvalgu VP16 aktivatsioonidomeenist (VP16-AD). Kolmas liitvalk sisaldas mutantset Gal4 aktivatsioonidomeni (Gal481-AD) ja LexA-BD. GAL481 on dominantne mutatsioon ning see mutant on konstitutiivselt aktiivne (st. ei vaja seostumist UASG järjestusele). Konstrueeriti ka kaks gal4 defektset pärmitüve.
Esimene neist sisaldas reportergeeni, mille promootoralas paiknes UASG järjestus. Teine sisaldas aga reporterkonstrukti, kus lacZ ekspressioon oli LexA OP kontrolli all. Liitvalgud ekspresseeriti gal4 defektsetes pärmitüvedes ja analüüsiti reportergeeni lacZ ekspressiooni. Katses kasutati ka metsiktüüpi tüve, kus Gal4 on funktsionaalne.
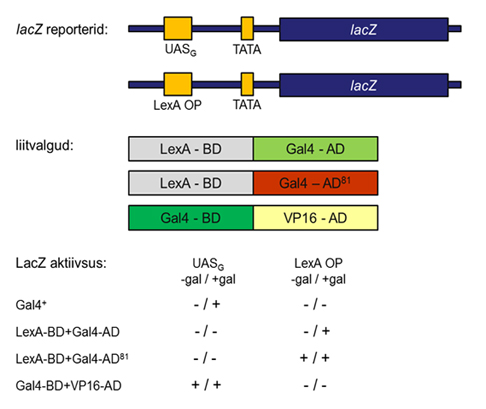
Joonis 5.
Gal4 domeeni vahetuse katse. Näidatud on kasutatud liitvalgud ja reportergeenid. Reportergeeni ekspresiooni analüüsiti galaktoosi puudumisel ja galaktoosi lisamisel.
Liitvalk, mis koosnes Gal4
aktivatsioonidomeenist ja LexA DNA-ga seostumise domeenist aktiveerib
reportergeeni ekspressiooni LexA OP järjestusele seostumisel ning glükoosi juuresolekul. Liitvalk
Gal4 DNA-ga seostumise domeen - VP16 aktivatsioonidomeen on aktiivne kui promootoralas on UASG järjestus. Galaktoos ei reguleeri selle liitvalgu aktiivsust. Liitvalk, mis sisaldas Gal481 aktivatsioonidomeeni ja LexA DNA-ga seostumise domeeni aktiveerib reportergeeni transkriptsiooni kui promootoris asub LexA OP järjestus. Galaktoos seda liitvalku ei reguleeri.
Nende katsete abil selgitati GAL geenide transkriptsiooni regulatsioon. Gal4 sarnane modulaarne organisatsioon on iseloomulik paljudele transkriptsioonifaktoritele.