Vastuselektsiooni võimaldavad markerid
Vastuselektsiooni e. negatiivset selektsiooni kasutatakse, et lahti saada tüves olevast markergeenist. See markergeen võib olla
- plasmiidi koosseisus. Sellisel juhul isoleeritakse rakud, kes on plasmiidi kaotanud.
- integreeritud kromosoomi. Sellisel juhul transformeeritakse rakkudesse markergeenist ülespoole ja allapoole jäävad järjestused ning selekteeritakse markergeeni välja rekombineerumist.
Pagaripärmis kasutatakse vastuselekteeritavate markeritena URA3, LYS2 ja CAN1 geene. Poolduvas pärmis leiavad vastuselektsioonil kasutamist ura4+ ja can1 geenid ning herpesviiruse tümidiin kinaas (hsv-tk+).
URA3 ja ura4+ on ortoloogid ning kodeerivad orotidiin-5'-fosfaat dekarboksülaasi, mis katalüüsib kuuendat reaktsiooni "de novo" pürimidiinide bisünteesi rajas:
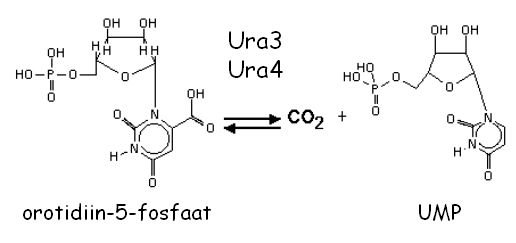
Mutatsioonid nendes geenides põhjustavad uratsiili auksotroofsust (Ura-). Mõlemaid markergeene on võimalik vastuselekteerida kasutades söötmes 5-fluoro-oroothapet (5-FOA). Selleks külvatakse rakud miinimumsöötmele, mis sisaldab uratsiili ja 0.1% 5-FOA. Kui rakud ekspresseerivad Ura3 või Ura4 valke, siis katalüüsitakse 5-FOA toksiliseks 5-fluorouratsiiliks ning rakud surevad.
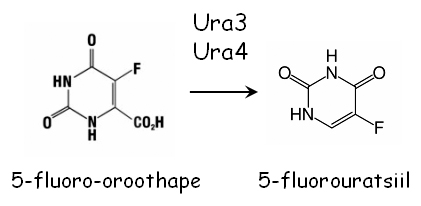
5-FOA-d sisaldaval söötmel jäävad ellu vaid ura3 ja ura4 mutandid. Selline negatiivne selektsioon töötab väga efektiivselt. Vähem kui 10-2 5-FOA resistentsetest kolooniatest on Ura+ fenotüübiga.
LYS2 (LYSine requiring) kodeerib  -aminoadipaat reduktaasi, mis katalüüsib viiendat reaktsiooni lüsiini biosünteesi rajas:
-aminoadipaat reduktaasi, mis katalüüsib viiendat reaktsiooni lüsiini biosünteesi rajas:
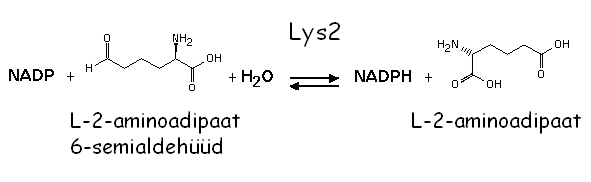
Mutatsioonid selles geenis põhjustavad lüsiini auksotroofsust - Lys-. Lys2 aktivatsiooniks on vajalik Lys5, mis muudab inaktiivse Lys2 apo-vormi katalüütiliselt aktiivseks holo-vormiks. Seepärast on nii lys2 kui ka lys5 mutandid sarnase fenotüübiga. Nendes mutantides koguneb toksiline vaheühend -aminoadipaat (-AA):
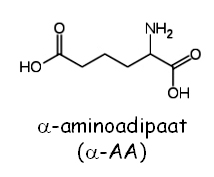
Mõlemad mutandid kasutavad -AA kui lämmastiku allikat. Prototroofsed tüved (st. Lys+) on selle toksilise ühendi suhtes tundlikud ning surevad. Lisaks plasmiidi vastuselektsioonile on võimalik eraldada ka spontaanselt tekkivaid lys2 mutante. Metsiktüüpi rakud plaaditakse söötmele, mis sialdab -AA-d. Sellel söötmel kasvavd üles vaid rakud, kus LYS2 ja harvemal juhul ka LYS5 geenides on tekkinud mutatsioon.
CAN1 (CANavanine resistance) kodeerib arginiini transporterit. Metsiktüüpi CAN1 geeni sisaldavad tüved on tundlikud toksilise arginiini analoogi kanavaniin sulfaadi suhtes:
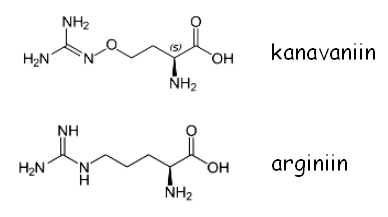
Kanavaniin transporditakse rakku arginiini transporteri Can1 poolt. Kui söötmes puudub arginiin, siis kanavaniin lülitatakse arginiini asemel peptiidi ahelasse ning see on rakkudele surmav. can1 mutatsioon muudab rakud kanavaniini suhtes resistentseteks. Diploidid CAN1/can1 on kanavaniini suhtes tundlikud. Kuna arg- mutandid vajavad arginiini permeaasi, et transportida söötmest rakku arginiini, siis can1 vastuselektsioonil ei saa kasutada arg- tüve. Lisaks arginiinile transpordib Can1 permeaas rakku ka histidiini. Seepärast ei soovitata vastuseleketsioonil kasutada ka his- mutante, sest can1 his- rakud on väga aeglase kasvuga.
Inimese herpesviiruse tümidiin kinaas (hsc-tk+): poolduva pärmi genoom ei kodeeri tümidiin kinaasi. Rakud, kuhu on sisse viidud tk+ geen ekspresseerivad funktsionaalset ensüümi. Tümidiin kinaasi üheks substraadiks on 5'-fluoro-2'-deoksüuridiin (FudR), mis fosforüleerituna on rakkudele toksiline:
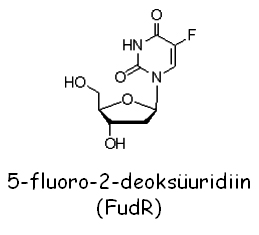
Seega rakud, kes ekspresseerivad hsv-tk+ on FudR tundlikud. Metsiktüüpi rakud, kellel puudub see markergeen, on FudR resistentsed.